H3K9メチル化とDNAメチル化による
転写抑制機構の全容の解明
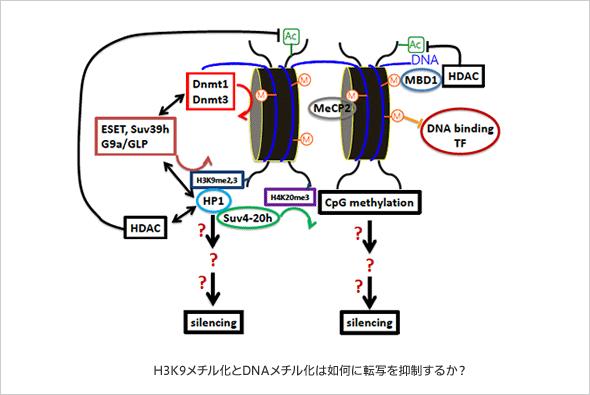
2003年のヒトゲノム計画の完了以降、様々な生物種の全ゲノムが解読されて来た。その結果明らかにされた共通する重要な知見の一つは、生物のゲノムは遺伝子よりもはるかに多くの転移因子に由来する配列によって占められているという点である。例えば、ヒトでは、遺伝子をコードする領域はわずか2%程度にしか過ぎないのに対して転移因子に由来する配列は50%以上にわたる。つまり、生命は転移因子とともに進化してきたとも言えるが、転移因子の転移はほとんどの場合宿主にとっては中立か不利益である。そのため、生物は転移因子に対して、様々な対抗する機構を備えてきた。一旦ゲノムに入り込んだ転移因子を抑える最初の機構は、転移因子の転写を抑制することにある。DNAのメチル化は転移因子の転写抑制に重要な役割を果たしており、植物や哺乳類を始めとする多くの動物ではDNAのメチル化が低下すると転移因子が活性化し転移が誘導されることが分かっている。しかし、DNAのメチル化が存在しない生物も存在する(例えば酵母)。近年の研究により、ヒストンH3の9番目のリジン残基(H3K9)のメチル化によるエピジェネティクス制御機構が転移因子の抑制に重要な役割を果たしていること、この機構はDNAのメチル化以上に生物種で広く使われている転写抑制機構であることが明らかとなった。これまでの解析から、H3K9ジメチル、トリメチル化修飾(H3K9me2,3)は転写抑制のエピゲノムマークとして機能し、このマークに親和性を示す共通した読み取り分子を呼び込むことが転写抑制のシグナル伝達に重要だと理解されている。実際、このマークが存在する生物種ではH3K9にメチル基を付加する酵素もH3K9me2,3に高親和性を示すクロモドメインをもつタンパク質であるHP1ファミリー分子も保存されている。しかし、HP1ファミリー分子がいかにして転写抑制を誘導するのか、はたまたHP1分子の呼び込みは転写抑制に(共通して)どのくらい重要なのか、実はよく分かっておらず、つまり“H3K9メチル化により誘導される転写抑制”の実体は十分に解明されていない。そこで、この種を超えて保存されているH3K9メチル化による転写抑制機構の全容を明らかにしたい。さらに、DNAのメチル化による転写抑制の実体に関しても、実はその全容はよくわかっていない。H3K9メチル化による制御と比較しながら、DNAメチル化による転写抑制の実体に関しても迫ってみたい。